2023
- Pib2 is a cysteine sensor involved in TORC1 activation in Saccharomyces cerevisiae
 Qingzhong Zeng, Yasuhiro Araki*, Takeshi Noda* Cell Reports プレスリリース
Qingzhong Zeng, Yasuhiro Araki*, Takeshi Noda* Cell Reports プレスリリース
- Characterization of Rab32- and Rab38-positive lysosome-related organelles in osteoclasts and macrophages Kazuya Noda, Shiou-Ling Lu , Siyu Chen , Kanako Tokuda , Yangjie Li , Feike Hao , Yoh Wada , Ge-Hong Sun-Wada , Shinya Murakami , Mitsunori Fukuda , Takashi Itoh, Takeshi Noda* Journal of Biological Chemistry Volume 299, Issue 10, October 2023, 105191 プレスリリース
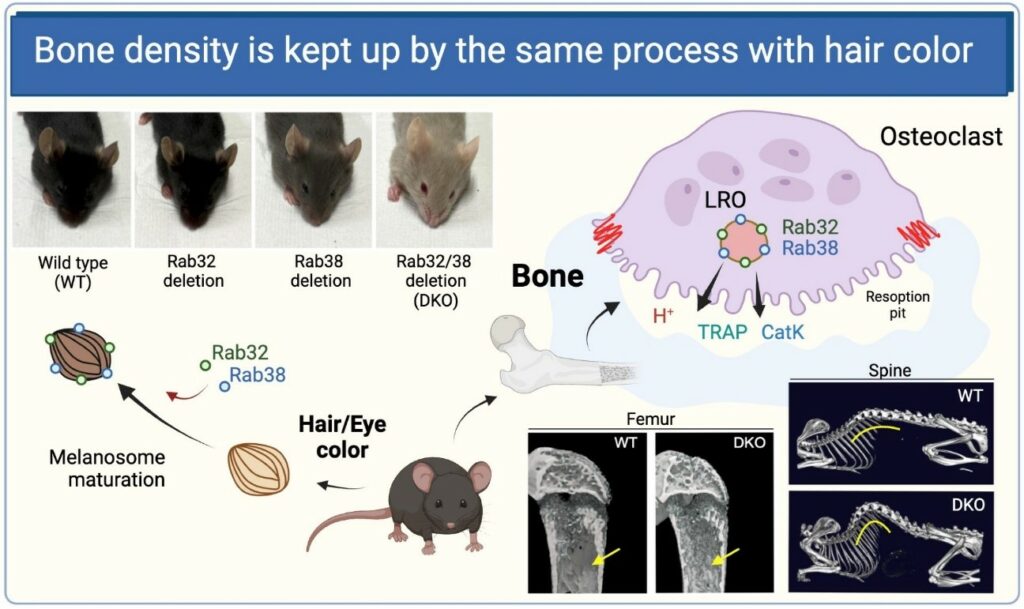
- Rab32 and Rab38 maintain bone homeostasis by regulating intracellular traffic in osteoclasts Kanako Tokuda, Shiou-Ling Lu*, Zidi Zhang, Yumiko Kato, Siyu Chen, Kazuya Noda*, Katsutoshi Hirose, Yu Usami, Narikazu Uzawa, Shinya Murakami, Satoru Toyosawa, Mitsunori Fukuda, Ge-Hong Sun-Wada, Yoh Wada, Takeshi Noda Cell Structure and Function 2023 Vol 48 2 p. 223-239 プレスリリース
- Opposing roles of RUBCN isoforms in autophagy and memory B cell generation CHAO-YUAN TSAI, SHUHEI SAKAKIBARA, YU-DIAO KUAN, HIROKO OMORI, MARUWA ALI EL HUSSIEN, DAISUKE OKUZAKI, SHIOULING LU, TAKESHI NODA, KEISUKE TABATA, SHUHEI NAKAMURA, TAMOTSU YOSHIMORI, HITOSHI KIKUTANI SCIENCE SIGNALING VOl 16 Issue 803
2022
- VEGF-Mediated Augmentation of Autophagic and Lysosomal Activity in Endothelial Cells Defends against Intracellular Streptococcus pyogenes Shiou-Ling Lu, Hiroko Omori, Yi Zhou, Yee-Shin Lin, Ching-Chuan Liu, Jiunn-Jong Wu, Takeshi Noda* mBio プレスリリース 日経バイオテク
- VEGF (vascular endothelial growth factor) provides antimicrobial effects via autophagy and lysosomal empowerment in endothelial cells Shiou-Ling Lu, Takeshi NODA Autophagy report Vol1 Issue 1 555-558
2021
- Vacuolar protein Tag1 and Atg1–Atg13 regulate autophagy termination during persistent starvation in S. cerevisiae Shintaro Kira, Masafumi Noguchi, Yasuhiro Araki, Yu Oikawa, Tamotsu Yoshimori, Aiko Miyahara, Takeshi Noda* Journal of Cell Science 134: jcs253682 Faculty opinion オートファジーフォーラム
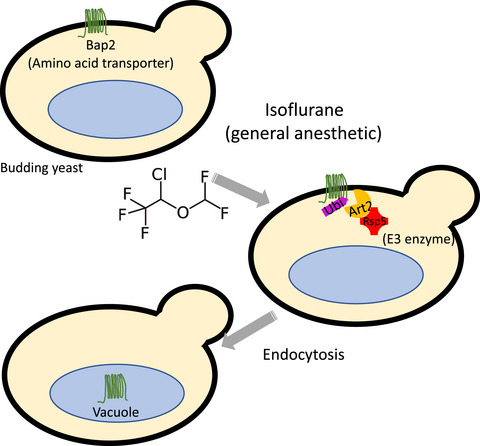
- Isoflurane induces Art2‐Rsp5–dependent endocytosis of Bap2 in yeast Fumi Kozu, Kanae Shirahama‐Noda, Yasuhiro Araki, Shintaro Kira, Hitoshi Niwa, Takeshi Noda* FEBS Open Bio 11, 11, 3090-3100
- A CRISPR/Cas9-based method for seamless N-terminal protein tagging in Saccharomyces cerevisiae. Shintaro Kira*, Takeshi Noda Yeast 38, 11 592-600
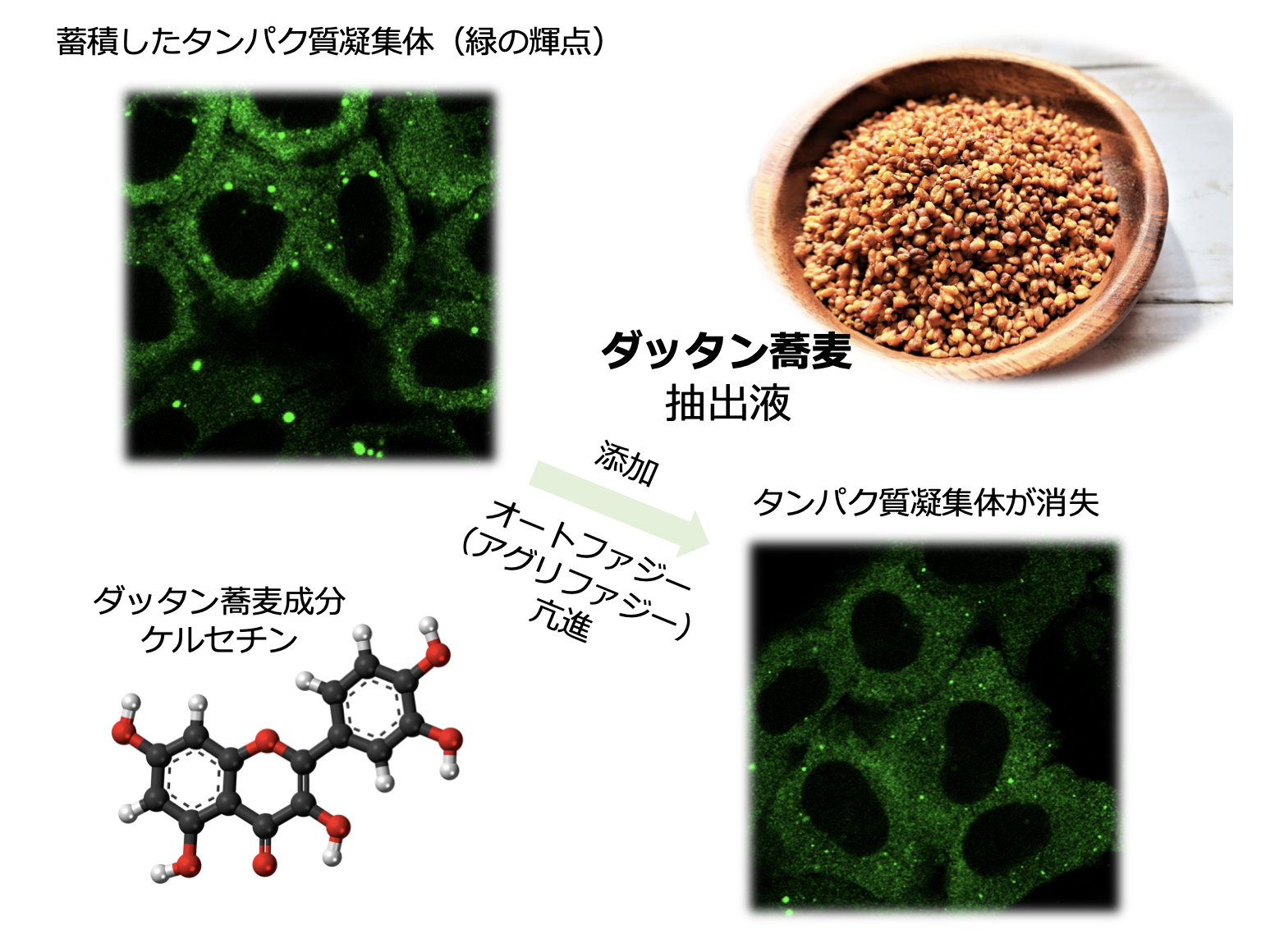
- Quercetin in Tartary Buckwheat Induces Autophagy against Protein Aggregations Sumiko Ikari, Qiang Yang, Shiou-Ling Lu, Yuancai Liu, Feike Hao, Guoqiang Tong, Shiguang Lu, Takeshi Noda* Antioxidants 10(8) 1217-1217 プレスリリース
- Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition).Daniel J Klionsky, Amal Kamal Abdel-Aziz, Sara Abdelfatah, Mahmoud Abdellatif, Asghar Abdoli, Steffen Abel, Hagai Abeliovic, Takeshi NODA, Yasuhiro Araki et al. Autophagy 1-382
2020
- Starvation-induced autophagy via calcium-dependent TFEB dephosphorylation is suppressed by Shigyakusan Sumiko Ikari, Shiou-Ling Lu, Feike Hao, Kenta Imai, Yasuhiro Araki, Yo-hei Yamamoto, Chao-Yuan Tsai , Yumi Nishiyama, Nobukazu Shitan, Tamotsu Yoshimori, Takanobu Otomo, Takeshi Noda* PLOS ONE 15(3) e0230156 プレスリリース

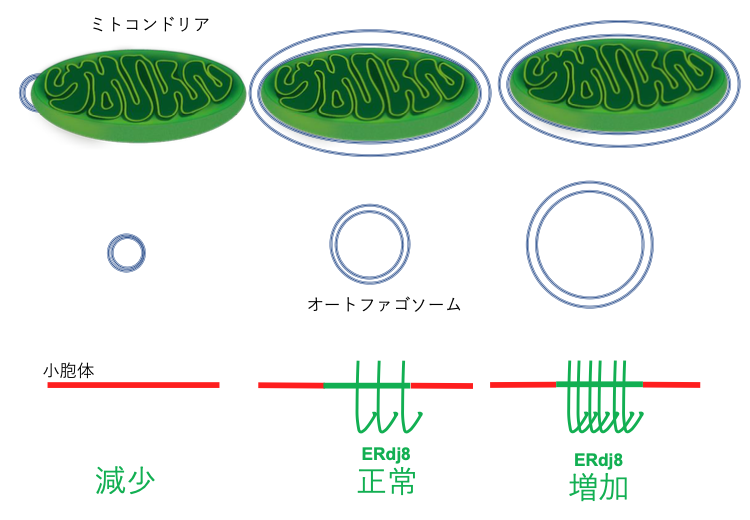
- ERdj8 governs the size of autophagosomes during the formation process. Yo-hei Yamamoto , Ayano Kasai, Hiroko Omori, Tomoe Takino, Munechika Sugihara, Tetsuo Umemoto, Maho Hamasaki, Tomohisa Hatta, Tohru Natsume, Richard I Morimoto, Ritsuko Arai, Satoshi Waguri, Miyuki Sato, Ken Sato, Shoshana Bar-Nun, Tamotsu Yoshimori, Takeshi Noda*, Kazuhiro Nagata* The Journal of Cell Biology 219 (8): e201903127 プレスリリース Editor”s choice in Science Academist Journal
- STEEP mediates STING ER exit and activation of signaling. Bao-Cun Zhang, Ramya Nandakumar, Line S Reinert, Jinrong Huang, Anders Laustsen, Zong-Liang Gao, Cheng-Long Sun, Søren Beck Jensen, Anne Troldborg, Sonia Assil, Martin F Berthelsen, Carsten Scavenius, Yan Zhang, Samuel J Windross, David Olagnier, Thaneas Prabakaran, Chiranjeevi Bodda, Ryo Narita, Yujia Cai, Cong-Gang Zhang, Harald Stenmark, Christine M Doucet, Takeshi Noda, Zheng Guo, Raphaela Goldbach-Mansky, Rune Hartmann, Zhijian J Chen, Jan J Enghild, Rasmus O Bak, Martin K Thomsen, Søren R Paludan Nature immunology 21(8) 868 – 879
- Nicotinamide Increases Intracellular NAD+ Content to Enhance Autophagy-Mediated Group A Streptococcal Clearance in Endothelial Cells. Cheng-Lu Hsieh, Shu-Ying Hsieh, Hsuan-Min Huang, Shiou-Ling Lu, Hiroko Omori, Po-Xing Zheng, Yen-Ning Ho, Yi-Lin Cheng, Yee-Shin Lin, Chuan Chiang-Ni, Pei-Jane Tsai, Shu-Ying Wang, Ching-Chuan Liu, Takeshi Noda, Jiunn-Jong Wu Frontiers in microbiology 11 117 – 117
- Autophagosome formation in relation to the endoplasmic reticulum Yo-hei Yamamoto, Takeshi Noda* Journal of Biomedical Science 27(1)
- オートファジーと相分離. 野田展生, 野田健司. 現代化学 増刊46. 35-38
- アミノ酸によるオートファジーの制御. 医学のあゆみ. 野田健司. 272. 9. 707-712
2019
- Group A Streptococcus Induces LAPosomes via SLO/β1 Integrin/NOX2/ROS Pathway in Endothelial Cells That Are Ineffective in Bacterial Killing and Suppress XenophagyYi-Lin Cheng, Chih-Feng Kuo, Shiou-Ling Lu, Omori Hiroko, Ya-Na Wu, Cheng-Lu Hsieh, Takeshi Noda, Shang-Rung Wu, Robert Anderson, Chiou-Feng Lin, Chia-Ling Chen, Jiunn-Jong Wu, Yee-Shin Lin mBio 0(5) e02148-19.
- Osteoblastic lysosome plays a central role in mineralization. Iwayama T, Okada T, Ueda T, Tomita K, Matsumoto S, Takedachi M, Wakisaka S, Noda T, Ogura T, Okano T, Fratzl P, Ogura T, Murakami S Science advances 5(7) eaax0672
- 栄養を感知して細胞成長を促すTORC1の制御機構. 荒木保弘, 野田健司.生産と技術. 2019. 71. 3. 77-80
2018
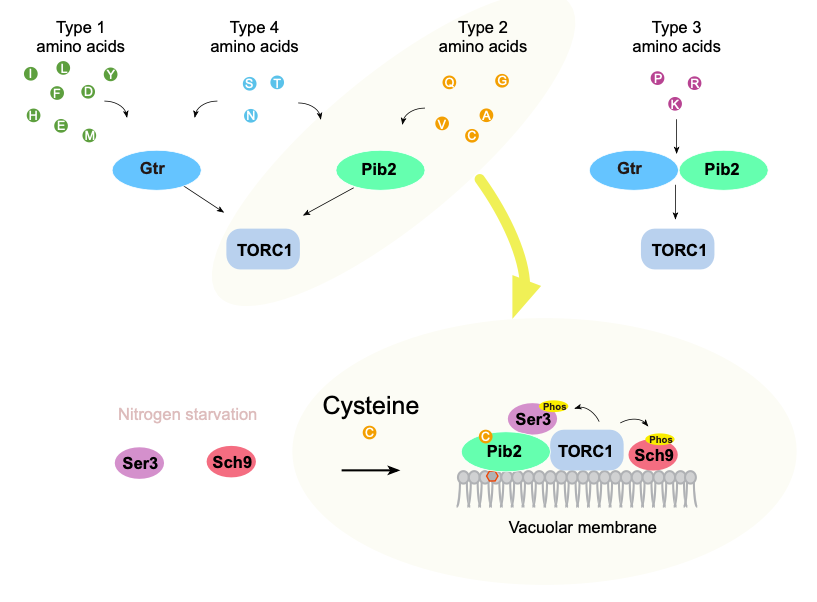
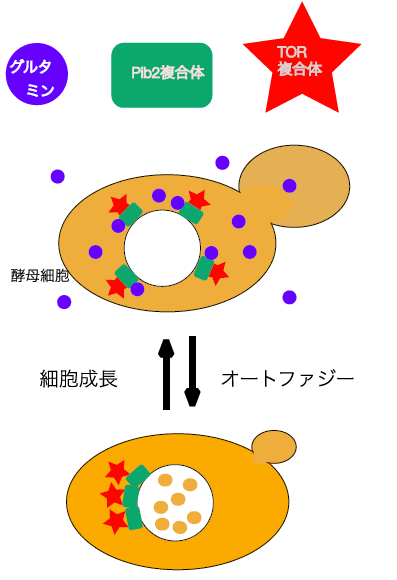
- Gtr/Ego-independent TORC1 activation is achieved through a glutamine-sensitive interaction with Pib2 on the vacuolar membraneUkai H, Araki Y*, Kira S, Oikawa Y, May A. I, Noda T* PLoS Genetics 14(4): e1007334. PI3P 結合タンパク質Pib2は、これまで知られていたGtr/Egoの経路と独立にTORC1を活性化することを見出した。Pib2とGtr を同時に消失させるとTORC1が液胞から遊離した。Pib2は特にグルタミンに反応してTORC1を活性化したが、グルタミン存在下に Pib2 とTORC1の相互作用が強まった。Pib2を含む複合体はグルタミンと直接相互作用した。これらのことからPib2を含む複合体が細胞内のグルタミンを感知してTORC1を活性化することを提唱した。プレスリリース 日本経済新聞電子版 日 経バイオテク
- Vacuole-mediated selective regulation of TORC1-Sch9 signaling following oxidative stressTakeda E, Jin N, Itakura E, Kira S, Kamada Y, Weisman LS, Noda T, Matsuura A* Mol Biol Cell Vol. 29, No. 4 377-522
- Rheb localized on the Golgi membrane activates lysosome-localized mTORC1 at the Golgi-lysosome contact siteHao F, Kondo K, Itoh T, Ikari S, Nada S, Okada M, Noda T*, J Cell Sci vol. 131 no. 3, jcs208017 mTORC1活性化因子の低分子量GTPase Rhebはこれまでリソソームに局在するモデルが提唱されてきたが、実際は主にゴルジ体に局在 し、かつRhebやmTORC1をゴルジ体に人為的に局在させても、その活性を維持することを見出した。さらにゴルジ体とmTORC1が物理的に接触するGolgi lysosome contact sitegが存在することを見出し、それらの異なるオルガネラ間の接触がmTORC1 の活性調節に関わる可能性を提唱した。Fuculty opinion 1 2
- Induction of selective autophagy in cells replicating hepatitis C virus genome. Mori H, Fukuhara T, Ono C, Tamura T, Sato A, Fauzyah Y, Wada M, Okamoto T, Noda T, Yoshimori T, Matsuura Y The Journal of general virology 99(12) 1643 – 16577
2017
- Ole1, fatty acid desaturase, is required for Atg9 delivery and isolation membrane expansion during autophagy in Saccharomyces cerevisiaeOgasawara Y, Kira S (equally contributed), Noda T*, Yamamoto A* Biology open 6: 1. 35-40 脂肪酸不飽和化酵素Ole1の欠損によりオートファゴソーム形成が不全となり、その原因の少なくとも1つがAtg9の輸送欠損であること を示しました。
- Endothelial cells are intrinsically defective in xenophagy of Streptococcus pyogenes Lu S, Kawabata T, Cheng Y, Omori H, Hamasaki M, Kusaba T, Iwamoto R, Arimoto H, Noda T, Lin Y, Yoshimori T* PLOS pathogens 13(7): e1006444/
- Microarray analysis of macrophage response to infection with Streptococcus oralis reveals the immunosuppressive effect of hydrogen peroxideMatsushima H, Kumagai Y, Vandenbon A, Kataoka H, Kadena M, Fukamachi H, Arimoto T, Morisaki H, Fujiwara N, Okahashi N, Kuwata H*. Biochem Biophys Res Commun 485(2):461-467
- Streptococcus sanguinis induces neutrophil cell death by production of hydrogen peroxide. Sumioka R, Nakata M*, Okahashi N, Li Y, Wada S, Yamaguchi M, Sumitomo T, Hayashi M, Kawabata S PLoS One 12(2):e0172223
- Autophagy in the context of the cellular membrane-trafficking system: the enigma of Atg9 vesiclesNoda T* Biochemical Society Transactions 45(6)1323-1331
- Regulation of Autophagy through TORC1 and mTORC1 Noda T* Biomolecues 7(3), 52
- Quantitative assay of macroautophagy using Pho8∆60 assay and GFP-cleavage assay in yeast Araki Y, Kira S, Noda T* Methods in Enzymology 588: 307-321
- オートファジーの機構の解明-駒場で芽吹き蕾を育み、岡崎で花開き、世界で果実を共有する 野田健司 科学 87. 1, 68-70
- 大隅良典先生のオートファジー研究:辺境から生まれたノーベル賞 野田健司 学術の動向 22 ,2 13-17
- 酵母が切り開いたオートファジー研究の曙 野田健司 オートファジー 南山堂 ISBN 978-4-525-13481-5
- オートファジーの誘導と抑制をつかさどる2つのキナーゼAtg1/Ulk1とTORC1荒木保弘 野田健司 オートファジー 南山堂 ISBN 978-4-525-13481-5
2016
- Atg9A trafficking through the recycling endosomes is required for autophagosome formation.Imai K, Hao F, Fujita N, Tsuji Y, Oe Y, Araki Y, Hamasaki M, Noda T*, Yoshimori T* Journal of Cell Science 120(20):3781-3791我々が発見したオートファジーに必要なタンパク質Atg9は膜貫通タンパク質である。今回哺乳類Atg9AのN末端の細胞質側突出部位に、クラスリンアダプターAP-2との結合部位を見出し、その部位に変異を導入したAtg9Aはリサイクリングエンドソームにとどまり、ゴルジ体へ戻れなかった。またTRAPPIIIのノックダウンでもリサイクリングエンドソームに蓄積した。このことからエンドソームから ゴルジ体へのAtg9の輸送がオートファジーに必須であることが判明した。このことは以前私達が、酵母で報告したモデルを哺乳類でも支持するものである In This Issue
- Streptococcus oralis Induces Lysosomal Impairment of Macrophages via Bacterial Hydrogen Peroxide. Okahashi N*, Nakata M, Kuwata H, Kwabata S Infection Immunity 84(7):2042-2050
- The PtdIns3-phosphatase MTMR3 interacts with mTORC1 and suppresses its activity Hao F, Itoh T, Morita E, Shirahama-Noda K, Yoshimori T, Noda T* FEBS Letters 590(1):161-173以前私たちはPtdIns3PのホスファターゼMTMR3の不活性型を発現させると哺乳類培養細胞で栄養飢餓などの誘導刺激がなくても オートファジーが誘導される現象を報告した。今回MTMR3がmTORC1と結合し、その活性を抑制するという現象を発見した。MTMR3不活性型によるオートファジー誘導は局所的なPtdIns3P濃度を上昇させること、およびmTORC1活性を抑制することがかかわることが示唆された。
- Differing susceptibility to autophagic degradation of two LC3-binding proteins: SQSTM1/p62 and TBC1D25/OATL1 Hirano S, Uemura T, Annoh H, Fujita N, Waguri S, Itoh T*, Fukuda M* Autophagy 12(2):312-326
- Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition) Kilonsky DJ* et ai, Noda T, Autophagy 2;12(1):1-222.
- オー トファジー: 長年の謎を解き明かした大隅良典博士と酵母 荒木保弘、野田健司 化学 71, 12, 19-22
- オー トファジー野田健司メンブレントラフィック 化学同人
- 免疫系のしくみ 免疫学入門 第4版Sompayrac L著 桑田啓貴 岡橋暢夫 訳
2015
- Dynamic relocation of the TORC1–Gtr1/2–Ego1/2/3 complex is regulated by Gtr1 and Gtr2Kira S, Kumano Y, Ukai H, Takeda E, Matsuura A, Noda T* Molecular Biology of the Cell 27(2):382-396オートファジーを制御するTORC1の上流ではGtr1とGtr2のヘテロ二量体のGTP結合タンパク質が機能する。Gtrタンパ ク質 はEgoというタンパク質複合体により液胞膜へ係留されているが、今回私たちはEgoの新しいサブユニットEgo2を同定した。Egoの 他のサブユニットの欠損では、Gtrに依存した TORC1の活性化が完全に失われるが、Ego2の欠損株では部分的にしか失われない。このことはEgo2の欠損株ではGtrの液胞膜局在が部分的に残っていることと相関 する。TORC1、Gtr、Egoの複合体は液胞膜上に均一に局在するがそれに加えて液胞膜上で凝集した局在をする。Gtr1が GTP型 をとるとき、TORC1、Gtr、Egoは液胞膜上に均一なものが増加し、Gtr2がGDP型をとる時、凝集した局在が優勢となった。凝集する場合、Gtr2がTORC1に直接結合することが寄与していることも明らかにした。このようにGtr1とGtr2がそれぞれ GTP/GDP型を変換することでTORC1活性とともにその局在を変化させることを明 らかにした。その生理的意義も議論している。オートファジーフォーラム
- Rabin8 suppresses autophagosome formation independently of its guanine nucleotide-exchange activity towards Rab8 Amagai Y, Itoh T, Fukuda M, Mizuno K* The Journal of Biochemistry 158(2):139-153
2014
- Reciprocal conversion of Gtr1 and Gtr2 nucleotide-binding states by Npr2-Npr3 inactivates TORC1 and induces autophagyKira S, Tabata K, Shirahama-Noda K, Nozoe A, Yoshimori T, Noda T* Autophagy 10(9) 54-67オートファジーの制御機構を理解するため、酵母ゲノムワイド変異株コレクションを栄養飢餓に置き、網羅的にオートファジーの活性をALP 法により測定した。今回その中でオートファジー活性を著明に低下させたNpr2およびNpr3の変異株に注目した。その哺乳類ホモログのノックダウン でも同様にオートファジーの誘導に欠損がみられた。我々はTOR活性がオートファジー制御の中心を担うことを発見してきたが、Npr2およびNpr3は低分子量 G タンパク質Gtr1の上流でTORC1の活性調節を介してオートファジーを制御していた。npr2変異株はGtr1のGTP結合型変異株と相同な表現型を示し、Gtr1のGTP加水分解に関わることが示唆された。さらに、Gtr1と二量体を形成する低分子量Gタンパク質 Gtr2が、TORC1に直接結合することでその活性を抑制していることを見出した。このように二つの機構によりTORC1が不活性 化し オートファジーが誘導されることが明らかとなった。オートファジーフォーラム
- Cell wall-anchored nuclease of Streptococcus sanguinis contributes to escape from neutrophil extracellular trap-mediated bacteriocidal activity Morita C, Sumioka R (eaually contributed), Nakata M*, Okahashi N, Wada S, Yamashiro T, Hayashi M, Hamada S, Sumitomo T, Kawabata S PLOS ONE 9(8):e103125
- Porphyromonas gingivalis promotes invasion of oral squamous cell carcinoma through induction of proMMP9 and its activation. Inaba H*, Sugita H, Kuboniwa M, Iwai S, Hamada M, Noda T, Morisaki I, Lamont RJ, Amano A Cellular Microbiology 16(1) 131-145P. gingivalis感染により、口腔癌細胞株SAS細胞から分泌されたMMP9前駆体が活性化され るととも に、口腔癌浸潤能が促進された。さらに、PAR2の活性化とともにERK1/2とp38のリン酸ならびに下流に存在するEts1と HSP27の活性化が認められ、NF-kB の核移行が観察された。一方、P. gingivalis ジンジパイン変異株の感染では、PAR2が活性化されず、MMP9前駆体の発現誘導ならびに癌浸潤能も促進されなかった。これらのことから、P. gingivalisのジンジパインがMMP9前駆体発現を誘導し、さらに細胞外に分泌された前駆体MMP9を活 性化し、口腔癌の浸潤を促進していることが示唆された。
- Hydrogen peroxide contributes to the epithelial cell death induced by the oral mitis group of streptococci Okahashi N*, Sumitomo T, Nakata M, Sakurai A, Kuwata H, Kawabata S PLOS ONE 9(1) e88136以前、我々は、ミティス群口腔レンサ球菌の産生する過酸化水素がマクロファージに対して細胞傷害性を示すことを報告した。そこ で、 ミティス群レンサ球菌の細胞毒性を様々な細胞について検討したところ、上皮細胞が過酸化水素に対する感受性を有し、細胞死に加えてIL-6 産生などの炎症応答も起こすことを見出した。これらの結果は、過酸化水素がミティス群レンサ球菌の病原因子のひとつであることを 示唆 している。
2013
- TRAPPIII is responsible for the vesicular transport from early endosomes to the Golgi apparatus that facilitates Atg9 cycling in autophagy.Shirahama-Noda K, Kira S, Yoshimori T, Noda T* Journal of Cell Science 126 4963-4973 TRAPPIII複合体が低分子量Gタンパク質Ypt1(Rab1ホモログ)の活性化因子GEFとして、初期エンドソームからゴルジ体への逆行輸送のプロセスで機能することを発見した。その経路は我々が発 見したオートファジーに必要な膜タンパク質Atg9の輸送に必要であったことから、これまでその実体が不明であった細胞内を動き回る多数のAtg9陽性の小胞は、エンドソームゴルジ体間をシャトルする輸送小胞であり、Atg9はそのような方法で細胞内にプールされているというモデルを示した。F1000 Prime
- Recruitment of the autophagic machinery to endosomes during infection is mediated by ubiquitin.Fujita N, Morita E, (equally contributed) Itoh T, Tanaka A, Nakaoka M, Osada Y, Umemoto T,Saitoh T, Nakatogawa H, Kobayashi S, Haraguchi T, Guan JL, Iwai K, Tokunaga F, Saito K,shibashi K, Akira S, Fukuda M, Noda T*, Yoshimori T* The Journal of Cell Biology 203(1) 115-128 細胞内に侵入したサルモネラが、それを取り囲む膜を障害した場合、膜タンパク質がユビキチン化される。そのときユビキチンに直接 Atg16Lが結合する等の機構により、オートファジーの分子装置がその周囲へ連れてこられることにより、オートファゴソーム膜が形成されることを示した。In This Issue F1000 Prime
- Hydrogen peroxide produced by oral streptococci induces macrophage cell death. Okahashi N*, Nakata M, Sumitomo T, Terao Y, Kawabata S PLOS ONE 8 e62563.ミティス群口腔レンサ球菌を in vitroでマクロファージに感染させると細胞死を誘発する。細胞死を引き起こす病原因子を調べたところ、菌が産生する過酸化水素が細胞死を誘導するという意外な事実が明 らかになった。
- Autophagy sequesters damaged lysosomes to control lysosomal biogenesis and kidney injury. Maejima I, Takahashi A, Omori H, Kimura T, Takabatake Y, Saitoh T,Yamamoto A, Hamasaki M, Noda T, Isaka Y, Yoshimori T* The EMBO journal 2336-2347
- Autophagosomes form at ER-mitochondria contact sites. Hamasaki M, Furuta N, (equally contributed), Matsuda A, Nezu A, Yamamoto A,Fujita N, Oomori H, Noda T, Haraguchi T, Hiraoka Y, Amano A*, Yoshimori T* Nature 495 389-393
- Osteoprotegerin-deficient male mice as a model for severe alveolar bone loss: comparison with RANKL-overexpressing transgenic male mice. Koide M., Kobayashi Y, Ninomiya T, Nakamura M, Yasuda H, Arai Y, Okahashi N, Yoshinari N, Takahashi N, Udagawa N* Endocrinology 154 773-782.
- The transcription factors Atf1 and Pcr1 are essential for transcriptional induction of the extracellular maltase Agl1 in fission yeast. Kato H, Kira S, Kawamukai M* PLoS ONE 8(11): e80572
- Morphological analysis of autophagy. Tabata K, Hayashi-Nishino M, Noda T, Yamamoto A, Yoshimori T*
Methods in molecular biology (Clifton, N.J.) 931 449-466
- 自己分解系としてのオートファジー制御の課題 野田健司, 吉良新太郎 実験医学 31(9) 1368-1373
- オートファジーによる病原菌捕獲の仕組み 野田健司 大阪大学歯学雑誌 57(2) 49-52
2012
- Identification of signaling pathways mediating cell cycle arrest and apoptosis induced by Porphyromonas gingivalis in human trophoblasts. Inaba H*, Kuboniwa M, Sugita H, Lamont RJ , Amano A. Infection and Immunity 80(8) 2847-2857.P. gingivalisが胎盤のヒト栄養膜細胞に感染するとDNA損傷シグナルであるATR、Chk2 およ び p53をリン酸化、ならびにG1期の停止とアポトーシス誘導中にp53の蓄積とアポトーシスの誘発に関連する転写因子Ets1の増加が認 められた。P. gingivalisの付着・侵入は、細胞周期G1期停止やアポトーシスを導くDNA損傷シ グナ ルの 活性に関与していることが示唆された。
- Porphyromonas gingivalis SerB mediated dephosphorylation of host cell cofilin modulates invasion efficiency. Moffatt C, Inaba H, Hirano T, Lamont RJ*. Cellular Microbiology 14(4) 577-588
- Distribution and molecular characterization of Porphyromonas gulae carrying a new fimA genotype. Yamasaki Y, Nomura R, Nakano K*, Inaba H, Kuboniwa M, Shirai M, Kato Y, Murakami M, Naka S, Matsumoto-Nakano M, Ooshima T, Amano A, Asai F. Veterinary Microbiology 161(1-2) 196-205.
- Guidelines for the use and interpretation of assays for monitoring autophagy. D. J. Klionsky*, 他(T. Noda 1270人中809番目) Autophagy 8 445-544
- Three-axis model for Atg recruitment in autophagy against Salmonella Noda T*, Kageyama S, Fujita N, Yoshimori T International Journal of Cell Biology vol. 2012, Article ID 389562
- オートファジーのTorとの邂逅 野田健司 細胞工学 31(12) 1323-1324